
植物に開花をもたらすものとは一体何なのでしょうか?この問いに対する完全な答えはまだ得られていませんが、開花を引き起こすメカニズムについては多くのことが明らかになっています。開花をもたらす要因はたったひとつではなく、マジカルな開花ホルモンが存在しているわけでもありません。植物はいくつかの刺激に反応して、さまざまな生理的・遺伝的な反応が連鎖的に引き起こされると最終的に茎頂部に花芽(かが・はなめ)が形成されます。主な要因のひとつが『光周性』とよばれる光の影響です。
光周性とは、植物が光の長さや光の色に反応して起こす生理的な変化のことです。人間や動物が目で光を感じるのとは違い、植物は光を『見る』ことはできません。私たちが光と認識する電磁波は、植物にとって生長サイクルを変化させたり、エネルギーを作り出したりするスイッチのような役割を果たします。一方、動物は光を使って周囲の世界を視覚的にとらえます。光には、『粒子(フォトン)』と『波』という二重の性質があります。周波数が高い(波長が短い)ほど、フォトンのエネルギーは強くなります(図1参照)。植物は特定の光の周波数をとらえ、そのエネルギーを利用して必要な化学反応を引き起こすしくみを持っています。
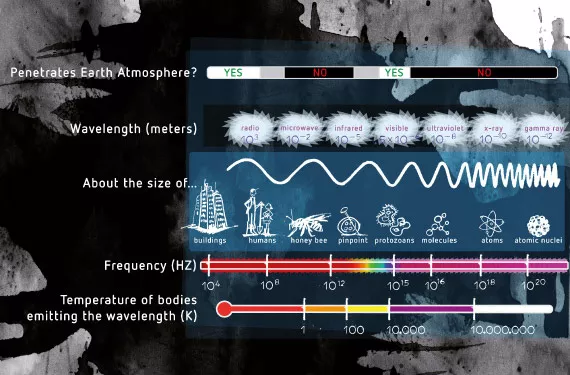
光スペクトルの色
植物が光エネルギーを必要とする理由は、大きく分けて2つあります。ひとつめは炭水化物を作るため、ふたつめは植物の細胞内で起こるさまざまなプロセスをコントロールするためです。ここではプロセスのコントロールについて説明しますが、そのために必要な光の範囲は、炭水化物を作る際に使われる光の範囲とほぼ同じです。植物が利用する光スペクトルは以下の基本的な4色です:
- UV (紫外線) 340 – 400 nm(ナノメーター)
- 青色光線 400 – 500 nm
- 赤色線 600 – 700 nm
- 遠赤色線 (赤外線の始まり) 700 – 800 nm
光の受容システム
実際の光スペクトルは、波長間で色がグラデーション状態で重なり合っているので波長ごとに境目がハッキリ分かれているわけではありません。植物は500~600nmの緑色光の一部も少しだけ利用しています。植物は異なる波長の光を吸収するためにさまざまな光受容色素(色を吸収する物質)を活用します。大まかに分類すると、植物の活動を制御する電磁エネルギーの4つの主要な波長域があり、それらは3つの光受容色素によってコントロールされています ;
- クリプトクロム(青色光・紫外線)
- フィトクロム(赤色光・遠赤色光)
- フォトトロピン(青色光・紫外線)
光の吸収点は、植物内の特定のプロセスをオン・オフするスイッチのように働き、その他のプロセスを調整する役割を持っています。人間は、光が物体に当たって反射された色(波長や周波数)を認識し、明るさが増したり減ったりすることでしか光の変化を感じません。しかし、植物は光の強さ(明るさ)だけでなく、波長の変化にも敏感です。
例えば、他の植物で影になって育つ植物は、青色光よりも赤色光や遠赤色光を多く受け取ります。また、日の出の際に赤色光から青色光へ変化する光のシフトや、日没の際に青色光から赤色光へと変化するシフトにも反応します。さらに、これらの変化が毎日どの時間に起こるかについても敏感に感じ取ります。植物のさまざまな色素(ピグメント)は、それぞれ特定の波長のエネルギーに反応し、異なる波長の比率によってスイッチが入るしくみになっています。また、光がないこと(暗やみ)さえも、植物の反応に影響します。これらの光の調節システムは、植物の開花というプロセスにも大きく関わっています。
光は、植物の自然なリズムを調整する重要な要素です。これは、光が動物の睡眠パターンに影響を与えるのと同じように、植物の生長や活動のタイミングを決定します。このような生物の体内リズムは「概日(がいじつ)リズム=サーカディアンリズム」と呼ばれ、すべての生命体に内在しています。植物にも1日の中でさまざまな活動があり、時間の変化に応じて活動が変わります。活動が活発な時間帯と休息する時間帯があり、エネルギーを消費するタイミングと生長や代謝がさかんになる時間帯が存在します。これらのプロセスは、だいたい24時間周期でくり返されるようにプログラムされています。
暗やみの中で光を吸収するための化学物質を生産するのは効率的ではありません(一部の物質は例外として生産されますが)。これは工場の生産ラインに例えることができます。適切なタイミングで必要なパーツが届き、在庫が管理され、最低限の材料が確保され、すべての部品がそろい、ようやく生産ラインが動きだすように、植物の物質生成は効率よくおこなわれます。光はこのリズムを決定する重要な要因であり、その存在だけでなく、波長や強度といった「質」も植物の活動に大きな影響を与えます。
植物は、受け取る光の量と質の両方を感知します。大気の状態や季節の変化などの環境要因によって、太陽光に含まれる色の構成は変化し植物はそれに適応します。植物が持つ特定の色素(ピグメント)が光の変化を感知し、他の外部要因や内部プロセスとのかねあいで、植物が「何をするか」「いつそれを行うか」が決まります。こうして、植物の体内時計が調整され、すべての生理的プロセスがバランスよく進行するのです。
植物が光の方向や強さを感知するのに重要な役割を果たすのがクリプトクロムという色素です。この色素は、植物の生長や生理的な反応を制御し、特に気孔の開閉、遺伝子の活性化、茎の伸長抑制、色素の合成、葉が太陽の動きに応じて向きを変える機能などを持ちます。
クリプトクロムは光の方向と強さを感知します。クリプトクロムは以下のような機能をコントロールしています;
- 気孔の開閉
- 遺伝子の転写と活性化
- 茎の伸びすぎを制御する
- 色素の合成
- および、太陽の動きに合わせて葉の向きを変える
もうひとつの青色光受容体であるフォトトロピンは、植物が光の方向に向かって動く性質(屈光性)を制御する役割を持っています。また、細胞内の葉緑体(クロロプラスト)の移動を調整し、光の強さに応じて配置を変えることで、過剰な光によるダメージを防ぐ働きをします。さらに気孔の開閉を調整するガードセル(孔辺細胞)の働きをうながす可能性があることも分かっています。

フィトクロム:PrおよびPfr
フィトクロムは、2種類の基本的な形態を持つ色素の複合体です:
- 赤色光に反応するタイプ(Pr)
- 遠赤色光に反応するタイプ(Pfr)
光を吸収する色素は、最も多く吸収する光の周波数を受け取ると活性化します(ただし、他の周波数や青色光も多少は影響を与えます)。これらの2つの色素は赤色光によってPrがPfrに変換され、その逆も然りというように、一般的に相互変換をくり返します(ただし、一部のPr/Pfrは、受ける光の量、光の強度、または光の質によっては再変換する能力を失う場合があります)。
植物の開花などの反応を引き起こす活性型はPfrです。赤色光は光形態形成(光が植物の発達に及ぼす影響)に最も大きな影響を与え、遠赤色光は時にPfrの作用を逆転させることがあります。
フィトクロムがコントロールする機能には以下のようなものがあります:
- 遺伝子の発現および抑制
- 遺伝子転写
- 幼苗や茎の伸長
- 発芽
- 光周性(開花反応)
- 日陰回避および光強度変化に対する適応
- クロロフィルの合成

赤色光への反応の例として、長日から短日への光周期の変化によって短日植物の開花がはじまる現象があります。これは、植物が 赤色光と遠赤色光(または無光)との比率の変化を感知すると、栄養生長(生長期)から生殖生長(開花期)へ移行するためです。植物が光を受けている間、PrとPfrの比率(Pr: Pfr)はほぼ同じに保たれ(実際にはPfrの方がわずかに多い)、赤色光によってPrがPfrに変換され、遠赤色光によってPfrがPrに戻ります。太陽が沈むと、遠赤色光の量が赤色光より増えるのでPrの量が増加します。するとPfrの濃度がわずかに高く、Prの濃度が低い状態になります。
暗さを感じた植物は、Prを生成し蓄積します。一方で、PfrはゆっくりとPrに分解され(その半減期は約2.5時間)。翌朝、再び太陽光があたるとPrとPfrの比率は元のバランスに戻ります。この過程を砂時計に例えると、Pfrは砂粒のようなものと考えることができます。現在の研究では、Pfrの濃度が低くPrが高い場合、短日植物が開花し、長日植物は開花しないことが分かっています。逆に、Pfrの濃度が高くPrが低いと、長日植物が開花し短日植物は開花しません。
長日と短日
例えば、昼間10時間 / 夜14時間で開花する植物(短日植物)と、昼間14時間/ 夜10時間で開花する植物(長日植物)があるとします。この場合、開花期に移行させる決定的な要因は「夜の長さ」 です(図3を参照)。短日植物 は、14時間の暗やみが必要 で、その間にPrが十分に蓄積されてPfrがPrに変換されることで、Pfrのレベルが十分に低くなる時間が長く続くと花芽が作られはじめます(開花期のスタート)。この変化は 一定の日数が経過すると不可逆的(元に戻らない) になります。長日植物では、このプロセスは基本的に逆の仕組みです。彼らは Pfrの濃度が高い状態に反応して開花します。

Pfrが優勢な時間が長くなると、植物は開花に移行します。しかし体内時計が整わないうちは(最初はそうであることが多い)、開花に移行するために必要な条件がすべてそろわず、体内時計が「追いつく」まで変化は起こりません。Pfrはフロリゲン(Florigen、開花シグナル)の抑制を解除するか、発現を促進し、そのシグナルによって植物は開花します。基本的に、Pfrの量が植物に夜の長さを知らせています。
フロリゲン — 開花のシグナル
フロリゲンはかつて仮説上のホルモンと考えられていましたが、現在ではFTmRNA として知られるメッセンジャーRNA であることが定説となりました。わかりやすく言えば、これは植物のDNAにある開花を促進する遺伝子領域「FT遺伝子」で生成されるタンパク質です。このタンパク質はカギのようなもので、特定のカギ穴を見つけるとピッタリはまります。そしてカギを回すと、次のプロセスに移るのです。
現在では、CO遺伝子という植物の光周性を抑制する遺伝子とフロリゲンが結びつくと、植物が生長期から開花期へと移行することがわかっています。つまり、植物の開花のメカニズムは、外部の環境からの合図によってコントロールされ、その影響で特定の遺伝子が働きはじめ、開花が進みます。植物は、光の変化を感知することで、これらすべてのプロセスを引き起こします。
植物の開花メカニズム
植物の開花メカニズムには、大きく分けて5つのタイプ があります。
- 短日植物(SDP:Short-Day Plants)短日・長夜条件で開花へ移行。
- 長日植物(LDP:Long-Day Plants)日・短夜条件で開花へ移行。
- 長日短日植物(LSDP:Long-Day Short-Day Plants)と..
- 短日長日植物(SDLP : Short-Day Long-Day Plants)...のそれぞれは、まず一定期間長日植物または短日植物の条件になってから、短日または長日になった条件で開花へ移行する。
- 最後に、同じ光の影響を受けるものの、日長以外の要因で開花へ移行する 日長中性植物(DNP)。
植物の開花は、受ける光の種類や光の質だけでなく、光がどれくらいの時間続くかも影響します(DNPs を除く)。
より正確に言えば、開花のスイッチが入るのは、夜の暗やみが続く時間で決まります。ただし、これは光が引き起こしたプロセスや代謝物(Pfr など)に基づいています。さらに、ここで説明したメカニズム以外にも、遺伝子やジベレリン酸(GA)などのホルモンとの相互作用を含む、さまざまな要因が開花に関わっています。
光はすべての生命にとって不可欠なものですが、特に植物にとっての光は、生長や代謝に必要な物質を作り出すだけでなく、日々のリズムや生活サイクルを整える役割も果たします。光は、植物の生存や繁殖に不可欠な要素を制御し、生命全体のリズムを決めるカギとなります。また、植物はすべての光に対して同じように反応するわけではありません。正常に生長するためには、適切な光のバランス(青色光と赤色光、赤色光と遠赤色光など)が必要です。さらに、光も過剰になれば、植物に悪影響を与えることがあります。とはいえ、光は植物にとって不可欠な存在ですが、それだけが生命活動に必要な要素のすべてではなく、生命を支えるひとつの要素に過ぎないことを覚えておいてください。
Bibliography
- Aushulz. "file:Chlorophyl_ab_spectra2.png." Wikipedia. http://en.wikipedia.org/wiki (accessed 4 10, 2011).
- Huang, T., Bōhlenius, H., Eriksson, S., Parcy, F., Nilsson, O. "The mRNA of the gene FT moves from leaf to shoot apex and induces flowering." Science 309 (2005): 1694-1696.
- Kasperbauer, M. J., and D. L. Karlen. "Plant Spacing and Reflected Far-Red Light Effects on Phytochrome-Regulated Photosynthate Allocation in Corn Seedlings." Crop Sci. 34 (1994): 1564-1569.
- Koning, Ross E. Photoperiodism. 1994. http://plantphys.info/plant_physiology/photoperiodism.shtml (accessed May 31, 2011).
- Taiz, L., and E. Zeiger. Plant Physiology. 3rd. Sunderland: Sinauer Associates, Inc., 2002.
- Zeevart, Jan A.D. "The Plant Cell 18:1783-1789 (2006)." The Plant Cell. August 2006. http://www.plantcell.org/content/18/8/1783.full#BIB18 (accessed June 9, 2011).



